A ATUAÇÃO DESTE COLEGIADO É REGIDA PELO REGIMENTO GERAL DE PÓS-GRADUAÇÃO DA UENF
Art. 15 - À Comissão Coordenadora do Programa compete:
I - demandar/sugerir as Disciplinas da área de concentração ou linha de pesquisa, bem como as do domínio conexo aprovadas pelo Laboratório e comunicar à SECACAD para cadastro;
II - estabelecer os requisitos específicos do Programa;
III - organizar instruções, normas ou regimentos específicos do Programa, além de planos e projetos a serem submetidos à apreciação da CPPG;
IV - informar à PROPPG os nomes dos professores credenciados e descredenciados, bem como o seu nível, segundo as regras estabelecidas pela CPPG, para homologação na CPPG;
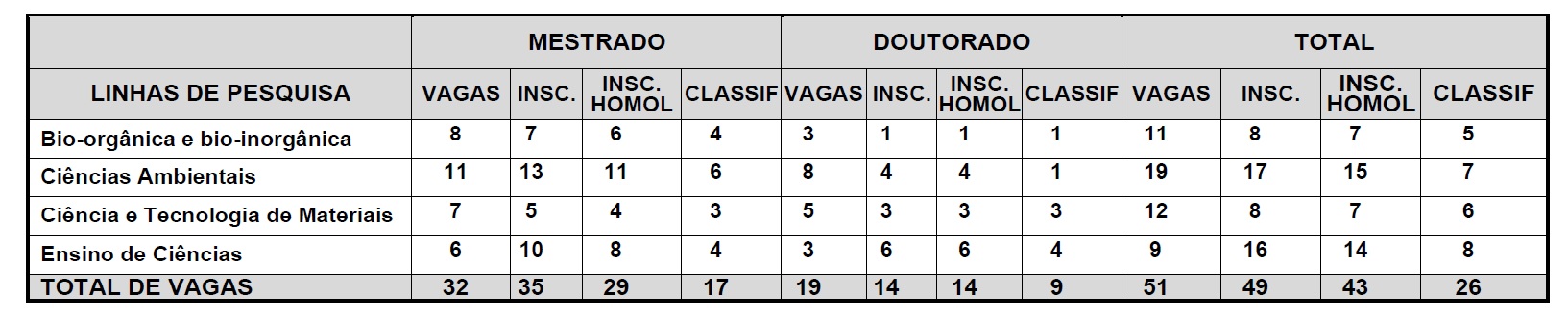
V - informar aos Centros pertinentes o número de vagas de Pós-Graduação do Programa para aprovação e encaminhamento à CPPG;
VI - coordenar a seleção de Candidatos qualificados para admissão no Programa, podendo designar para tal uma comissão especial constituída por professores credenciados pelo Programa;
VII - estabelecer normas para funcionamento de Seminários e indicar seu Coordenador;
VIII - aprovar a indicação de orientadores e, quando aplicável, os Planos de Estudos;
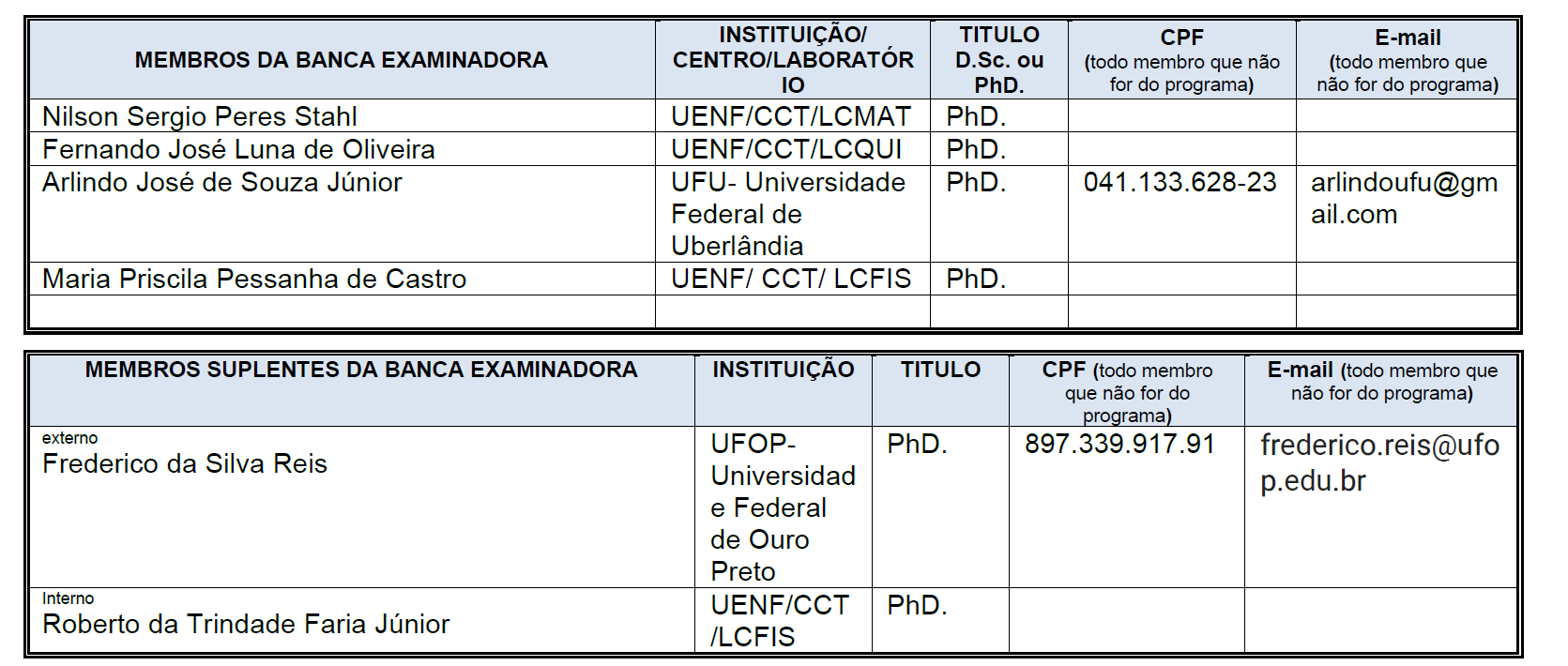
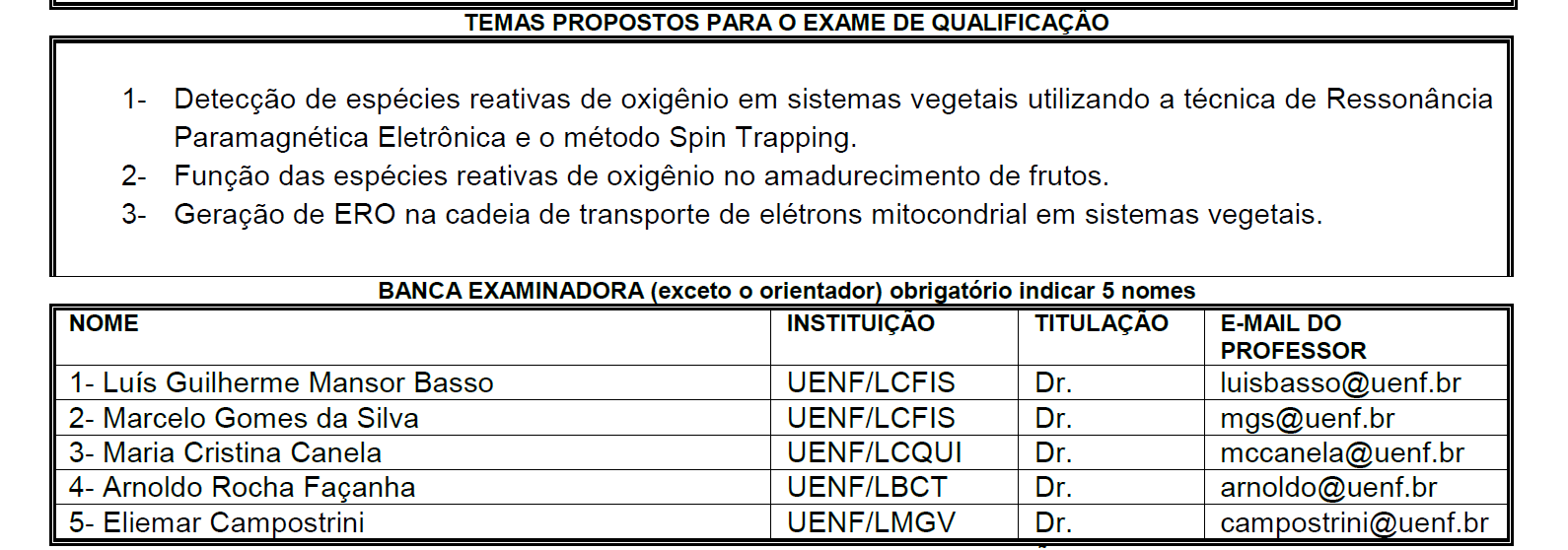
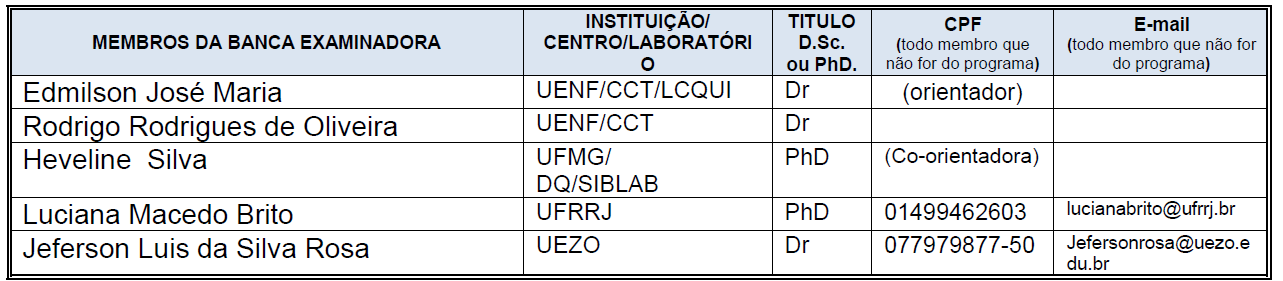
IX - aprovar a constituição de bancas de exame de projeto de dissertação ou de tese, de exame de qualificação e defesa de dissertação ou de tese;
X - propor à CPPG o desligamento de estudantes do Programa, por motivos acadêmicos ou disciplinares;
XI - indicar à CPPG os Candidatos selecionados ao Programa, destacando em ordem de classificação aqueles que poderão pleitear bolsas de estudo de acordo com as normas vigentes;
XII - apreciar ou propor convênios ou ajustes de cooperação de caráter acadêmico ou financeiro, para suporte ou desenvolvimento do Programa;
XIII - receber, apreciar, deliberar e encaminhar sugestões, reclamações, representações ou recursos, de estudantes ou professores, sobre qualquer assunto de natureza didático-científica, pertinentes ao Programa;
XIV - indicar à CPPG os Candidatos em condições de receber títulos de Pós-Graduação;
XV - atuar como órgão informativo e consultivo da CPPG;
XVI - deliberar sobre a prorrogação do prazo de conclusão da Pós-Graduação como previsto no CAPÍTULO I, Art. 3º e informar à SECACAD, em tempo hábil para viabilização da matrícula.
CAPITULO I
Art. 3º - O Mestrado terá duração mínima de 12 (doze) meses e máxima de 24 (vinte e quatro) meses e o Doutorado terá duração mínima de 24 (vinte e quatro) meses e máxima de 48 (quarenta e oito) meses contados a partir da data de admissão no Programa.
§ 1º- O pós-graduando poderá solicitar à Comissão Coordenadora do Programa (CCP) prorrogação por até mais 06 (seis) meses. Em sendo aprovada a solicitação pela CCP a mesma deverá informar à Pró-Reitoria de Pesquisa e Pós-Graduação (PROPPG) e à Secretaria Acadêmica (SECACAD).
§ 2º- Em casos excepcionais o pós-graduando poderá solicitar extensão da prorrogação por até 06 (seis) meses. Para tal deverá encaminhar a solicitação de extensão à CCP com uma antecedência de 03 (três) meses do vencimento do prazo, acompanhada dos seguintes documentos: justificativa da solicitação, parecer do orientador, relatório de atividades com respectivo cronograma e uma versão preliminar da dissertação ou da tese. O estudante deverá ter cumprido todas as exigências regimentais, exceto a defesa. A falta de um desses documentos impedirá sua apreciação pela CCP. A CCP após aprovação da solicitação encaminhará a mesma para avaliação da Câmara de Pesquisa e Pós-Graduação (CPPG).
Art. 16 - São Atribuições específicas do Coordenador:
I - convocar e presidir as reuniões da Comissão Coordenadora do Programa;
II - assinar, quando necessário, processos ou documentos submetidos ao julgamento da Comissão Coordenadora;
III - encaminhar os processos e deliberações da Comissão Coordenadora às autoridades competentes;
IV - promover entendimentos, com a finalidade de obter recursos humanos e materiais para suporte do desenvolvimento do Programa;
V - responder pelo Programa perante os Órgãos Superiores da UENF;
VI - zelar pela destinação dos recursos oriundos do Programa de Apoio à Pós-Graduação, da CAPES, e demais fontes financiadoras com mesmo fim;
VII - responder sobre o cumprimento das normas de concessão de bolsas de estudo;
VIII - disponibilizar informações aos estudantes sobre os prazos, normas e demais exigências, bem como sobre seus direitos e deveres;
IX - manter a SECACAD informada sobre a situação dos estudantes com relação aos respectivos orientadores, áreas de concentração e linhas de pesquisa.